

本文为双乙酰专题的最后一篇,主要探讨一些之前没有讨论的一些知识盲点和进阶工艺。
使用固定酵母连续发酵法减少双乙酰
在工业应用上,会用到连续发酵(Continuous fermentation)搭配固定酵母(Immobilized Yeast)工艺的方法减少双乙酰。
连续发酵
连续发酵法是20世纪70年代诞生的一种技术。最开始的目的就是为了更加高效率地生产乙醇。连续发酵,不同于单桶发酵技术,最大限度避免酵母的生长期,迟缓期和成熟期,而是直接进入发酵阶段,并且可以一直发酵而不停止。
其原理就是将培养基(也就是麦芽汁)以一定速率流入反应器内(也就是发酵罐),同时以固定速率流出发酵液。通过工业控制可以找到流入、流出与发酵率之间的最优解。
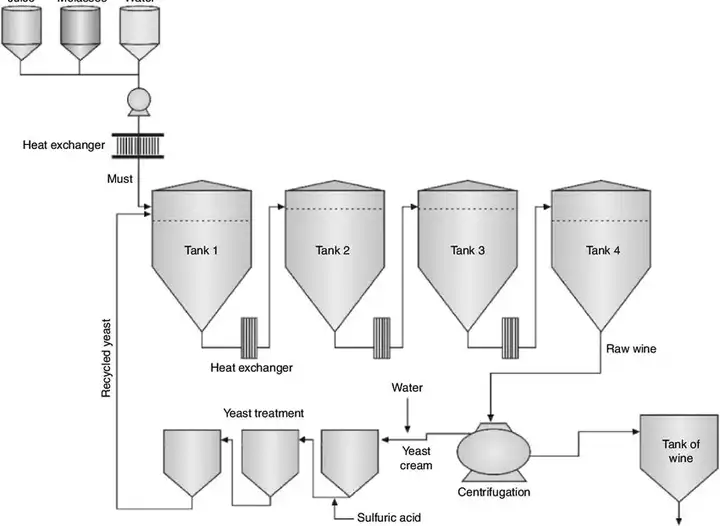
上图就为多罐连续发酵,将不同发酵度的发酵液经过多个发酵罐发酵。这样可以保持连续的发酵进程而不停止。当然也有单桶,下面会讲到。
尽管上面提到,可以找到流入/流出与发酵率之间的最优解。但传统的连续发酵工艺,发酵度不高依旧是个问题,其中一个解决方法就是多罐,但成本很高;且连续补充麦芽汁、桶于桶之间交换发酵液等容易导致染菌。所以就引入了固定化酵母这个工艺。
固定化酵母
固定化酵母的原理是,将酵母集中固定在载体上,让其进行快速繁殖,使载体上聚集高浓度的酵母细胞。流经这些高浓度酵母聚合体的麦芽汁,麦芽汁中的葡萄糖会被快速消耗并转化为乙醇(及一些副产物)。
固定酵母的方法,会有所不同。吸附法,使用谷物纤维(玉米芯、使用过的谷物壳)作为酵母载体,这类属于吸附;而最常用,包括家酿都可以使用的是包埋法,使用海藻酸钠(微溶于水)作为载体,进行酵母固定化。
被固定的酵母浓度很高,加上不断繁殖而游离出的酵母,发酵罐的酵母总浓度比非固定化酵母的批次更高。固定酵母的浓度可以去到非固定酵母浓度的1.5-2陪,所以反应速度更加迅速,某些研究表明发酵时间可以从几天减到一天甚至十几个小时;而高浓度的酵母有效抑制杂菌的滋生;加上收集方便,方便重复利用。
但被固定的酵母始终会受限于载体的面积大小,生长率有所下降,所以一般是经过培育后再进行使用。而固定酵母用于普通的酿酒中,始终不太适合,因为其快速的反应,容易导致其他副产物的失控。
目前比较常用的固定酵母连续发酵方法,是使用气升环流反应器(Gas Lift Reactor),如下图所示是一个双桶结构,一个用于发酵,一个用于成熟。R1反应器里塞满了被固定的酵母及其载体(载体为废弃谷物),底部充气,侧部进麦芽汁,气体压力让麦芽汁运动起来,在预先塞满了固定酵母里形成环流,避免某一区域酒精浓度过高导致发酵缓慢。

上图另外一个反应器,R3,也塞满了被固定的酵母及其载体(玉米芯),用于啤酒成熟。这里就没有使用环流,而是使用填充的方法进行成熟。
两个桶均为底部入原料,顶部出反应物,是因为当液位升高的时候,酒精比水轻,可判断最上层的液体已经经过了固定化酵母的发酵,所以顶部出产物。
那么为什么与双乙酰相关呢?其实R3里的啤酒成熟原理和普通的成熟差不多,双乙酰休止也是如此。不过由于酵母浓度更高,加上可以采用环流式反应器,使α-乙酰乳酸均匀被降解,所以效率会更高。
氧气影响究竟有多大?
其实在之前的搜寻资料中,很多资料会提到,过晚的充气(Aeration)会导致双乙酰的产生。所谓充气就是往麦芽汁里打入一定量的空气,目的是让酵母利用氧气进行繁殖,氧气有助于酵母合成甾醇,而甾醇正是合成细胞壁的重要物质。
氧促反应 or 非氧促反应
首先,过晚充气导致双乙酰过量的理论似乎和乙酸乳酰的非酶“氧促”反应有关。但第一篇中我就有提到,乙酰乳酸转化成双乙酰这个过程,是非酶反应,但是否跟氧气相关,我自己并没有给出定论。
在搜寻的论文,有两个观点。第一个,酿酒酵母基因库中给出的化合物合成降解通路中,双乙酰的前驱物,α-乙酰乳酸,转化为双乙酰的过程是一种自发生的反应,在合适的温度和pH下,会加快反应速度,并没有氧气参与反应。而双乙酰借助NADH还原为乙偶姻的反应,是不可逆的。这两种反应均不需要氧气的参与,“氧气导致双乙酰增加”是不成立的。
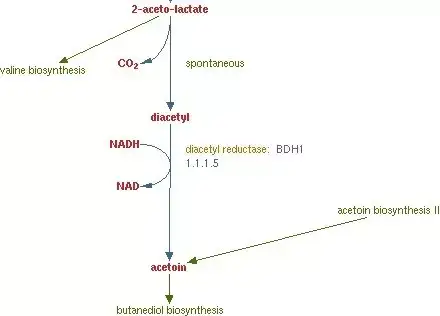
第二个观点,某些论文(尤其是关于乳酸乳球菌Lactococcus lactis,与酵母一样是兼性厌氧)在阐述双乙酰的生物生物合成的时候,会提到由α-乙酰乳酸转化为双乙酰的过程需要氧气参与,这些论文都集中在NCDO 763这个菌种。
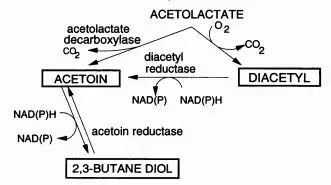
只不过,在KEGG基因库(京都基因與基因组百科全書)上查到的其他乳酸乳球菌的生物合成通路中,有关双乙酰的合成也是没有氧气参与。所以究竟后一观点是个例还是某些错误,不得而知。但起码在α-乙酰乳酸转化为双乙酰的反应,不用氧气是主流观点。
潜在的氧化反应?
而对于“氧气”在双乙酰含量增加的角色里的另一个假设是:双乙酰的还原产物乙偶姻,会不会重新被氧化回双乙酰呢?
比较可惜的是,通过乙偶姻氧化制备双乙酰,在现代工业上,会使用一些强氧化剂来反应。其中来自中国的两项制备双乙酰的专利,一个是使用三氧化铁与乙偶姻反应制备双乙酰;另一个使用二氧化锰、硫酸铜、硫酸铁等氧化剂,氧化乙偶姻制备双乙酰。似乎乙偶姻并不是有氧气就可以氧化为双乙酰。
氧气在其他阶段是否有影响?
而在一些实验中,证实了在成品啤酒中通入空气数日,都不会用双乙酰的增加。
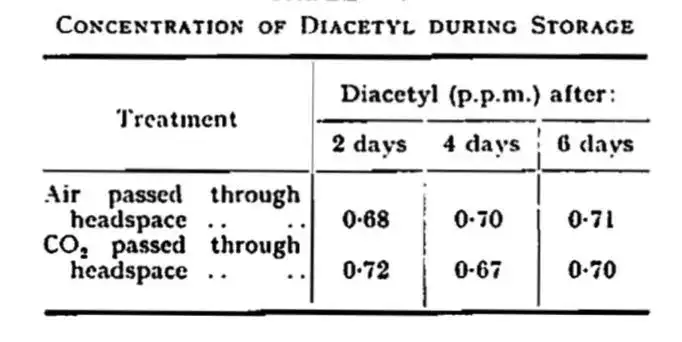
而且一些类似的实验,例如对发酵液长达48小时的通氧,然而发酵结束后,不同通气时间和量所产生的双乙酰都能够在限定的范围内。

其实还有一个较为极端的例子是开放式发酵(Open fermentation)。开放式发酵顾名思义就是发酵容器没有盖子,是接触着空气去发酵。这种发酵方法,主要用在德式小麦的酿造上。但并没有证据表明开放式发酵会带来双乙酰味道,同时德式小麦的标准风格也是不允许存在双乙酰味道。

那么究竟是什么原因大家会提及在发酵中通氧或充气会产生大量双乙酰?以下是我个人的推断。
首先综上所述,氧气对于双乙酰的产生并不是决定性因素。而个人认为在低温发酵的啤酒里,因为氧气原因导致双乙酰增加的可能性会更大。
在上一篇文章中,有提及一个参数,氨基酸的摄取率(Amino Acid Uptake Rate),在不同的酵母阶段有所不同,而缬氨酸(Valine)属于接种酵母后12个小时才会高速摄取的氨基酸。而双乙酰的残留很大程度上取决于α-乙酰乳酸流出细胞后自发反应成双乙酰,所以及时消耗α-乙酰乳酸用于生产缬氨酸成为减少双乙酰产生的关键。
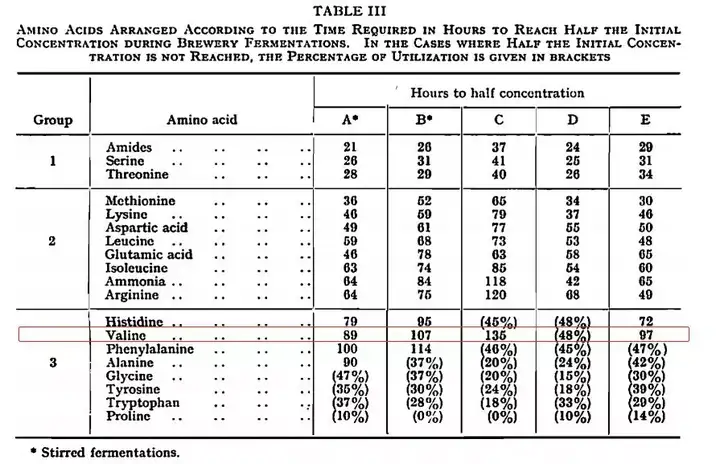
假如在发酵的更后段通入氧气,其实会促成酵母的有氧繁殖。大量的细胞繁殖,其实就等于回到第一阶段(生长阶段),这个阶段缬氨酸摄取率低,所以容易几率大量的α-乙酰乳酸,并进而转化为双乙酰。

而提到低温的原因在于,低温发酵啤酒往往比高温发酵更加谨慎地使用(甚至不用)双乙酰休止的过程(包括α-乙酰乳酸转化为双乙酰、双乙酰转化为乙偶姻两个过程),导致大量的α-乙酰乳酸潜伏于啤酒中,往后的运输及储藏过程就会因为升温而慢慢转化为双乙酰。
当然,对于已经储藏一段时间的啤酒中发现双乙酰,除了考虑储藏温度及工艺不对之外,还需要考虑一下是否有细菌污染,导致双乙酰超标。
麦芽与氨基酸
这里简单介绍一下各种谷物、麦芽制备、糖化、煮沸与FAN之间的关系,之后会做个专题。
麦芽拥有大量的氨基酸,用麦芽制备麦芽汁,而衡量其中的氨基酸含量,会用到FAN,Free Amino Nitrogen,游离氨基氮。关于FAN与双乙酰的关系在上一篇文章已经提及。所以在这里我只说麦芽的品种、制备及糖化以及煮沸对FAN的影响。
品种
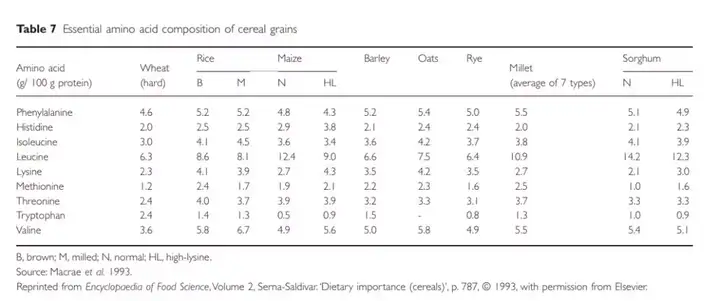
不同的谷物品种所含有的必要氨基酸含量不一样,下图给出了不同种类的谷物所含的必要氨基酸含量,可以看到缬氨酸含量最高是脱壳大米。
制备
麦芽制备是指对大麦/小麦等进行发芽培养,出芽后,进行干燥、烘干等工艺,最终获得可用于酿酒的麦芽。出芽的目的在于发芽过程,让大麦/小麦生成各种酶,这些酶在酿酒的糖化过程至关重要。
可以看到氨基酸含量在不同种类的大麦品种的制备过程,有不同的表现。但大致的趋势是发芽到96小时前基本是呈正增长。
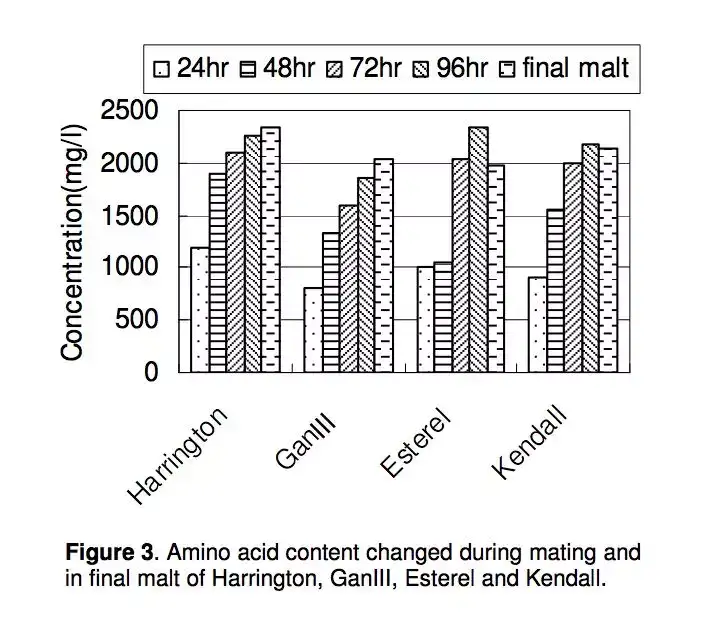
而对同一品种的麦芽(品种为Copeland)的不同制备方式,包括浸泡、出芽、烘烤过程的不同,对氨基酸的影响也不尽相同。而第三种制麦方式将会带出更多缬氨酸。
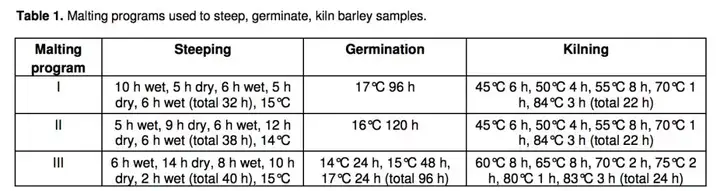

糖化
即使是比较大的酒厂,都很难说自己选择原材料去制备麦芽,时间、制作成本很不划算,所以也只能说是单纯地说下。但是糖化过程则是可控的。
以下是两种不同糖化温度对各种氨基酸的影响。55度的糖化温度(恰好是β-淀粉酶作用温度,尽管两者并没有联系),缬氨酸及其他氨基酸的浓度最高。

而不同的温度及pH值也会对氨基酸提取有影响。下图展示了在不同pH值、温度下氨基酸提取的量的不同。pH 5.2-5.4之间,缬氨酸的浓度最高。

煮沸
当然,煮沸过程也会伴随着氨基酸的流失,珠江啤酒厂与华农就做过相关的检测。但论文并未提到煮沸时间的长短,所以仅供参考。另据知情人士透露,珠江酒厂大约煮沸时间60分钟。
而煮沸时添加酒花也会导致氨基酸的浮动。

与麦芽相关的部分暂时介绍到这边。其实对于一般家酿爱好者而言,能够操控的就是糖化过程。
总结
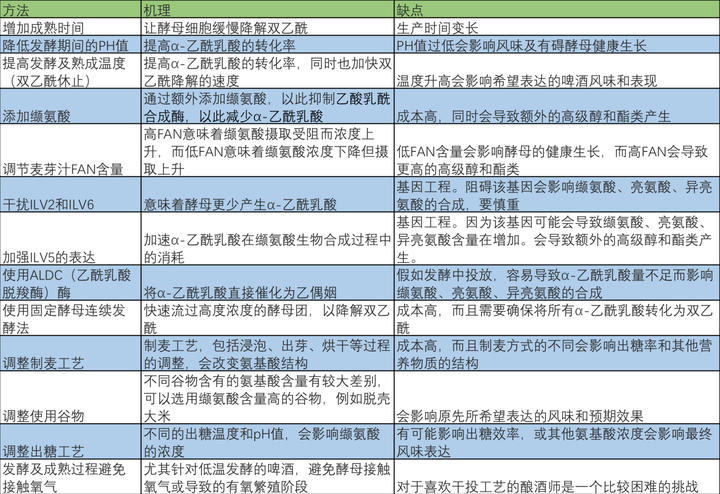
最后借鉴IBD(酿造与精馏研究所),用一张表为大家总结各个控制双乙酰的方法,从简单的双乙酰休止到基因工程的应用。
这个双乙酰专题到此结束,尽管还有很多的未知,且目前最简单的解决方法依然是做双乙酰休止。但知道多点其他原理并不是一件坏事。最后多谢各位朋友的大力支持,才有知识量如此庞大的文章。当然,也欢迎各位和我继续深入探讨有关双乙酰的问题。





